The following steps in the respiratory process are to release and utilise the energy stored in NADH+H+ and FADH2. This is accomplished when they are oxidised through the electron transport system and the electrons are passed on to O2 resulting in the formation of H2O. The metabolic pathway through which the electron passes from one carrier to another, is called the electron transport system (ETS) (Figure 14.4) and it is present in the inner mitochondrial membrane. Electrons from NADH produced in the mitochondrial matrix during citric acid cycle are oxidised by an NADH dehydrogenase (complex I), and electrons are then transferred to ubiquinone located within the inner membrane. Ubiquinone also receives reducing equivalents via FADH2 (complex II) that is generated during oxidation of succinate in the citric acid cycle. The reduced ubiquinone (ubiquinol) is then oxidised with the transfer of electrons to cytochrome c via cytochrome bc1 complex (complex III). Cytochrome c is a small protein attached to the outer surface of the inner membrane and acts as a mobile carrier for transfer of electrons between complex III and IV. Complex IV refers to cytochrome c oxidase complex containing cytochromes a and a3, and two copper centres.
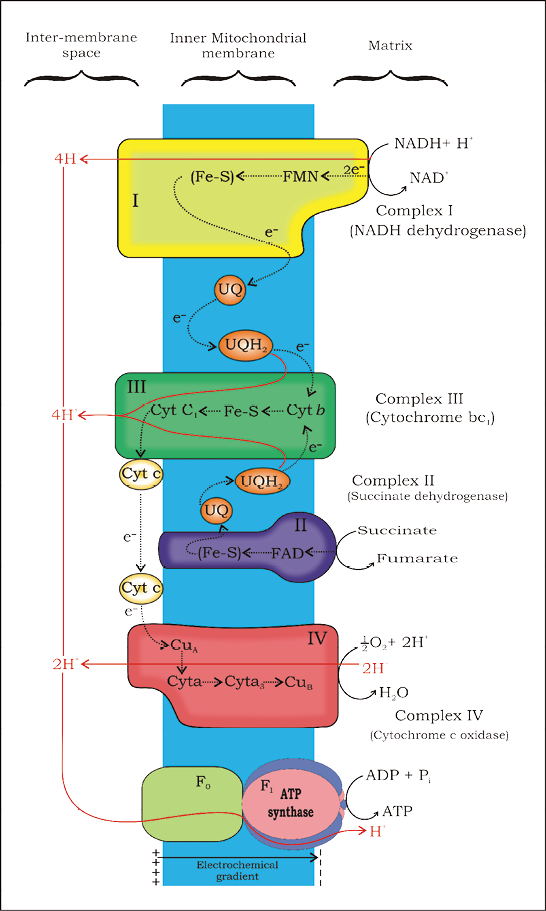
Figure 14.4 Electron Transport System (ETS)
When the electrons pass from one carrier to another via complex I to IV in the electron transport chain, they are coupled to ATP synthase (complex V) for the production of ATP from ADP and inorganic phosphate. The number of ATP molecules synthesised depends on the nature of the electron donor. Oxidation of one molecule of NADH gives rise to 3 molecules of ATP, while that of one molecule of FADH2 produces 2 molecules of ATP. Although the aerobic process of respiration takes place only in the presence of oxygen, the role of oxygen is limited to the terminal stage of the process. Yet, the presence of oxygen is vital, since it drives the whole process by removing hydrogen from the system. Oxygen acts as the final hydrogen acceptor. Unlike photophosphorylation where it is the light energy that is utilised for the production of proton gradient required for phosphorylation, in respiration it is the energy of oxidation-reduction utilised for the same process. It is for this reason that the process is called oxidative phosphorylation.
You have already studied about the mechanism of membrane-linked ATP synthesis as explained by chemiosmotic hypothesis in the earlier chapter. As mentioned earlier, the energy released during the electron transport system is utilised in synthesising ATP with the help of ATP synthase (complex V). This complex consists of two major components, F1 and F0 (Figure 14.5). The F1 headpiece is a peripheral membrane protein complex and contains the site for synthesis of ATP from ADP and inorganic phosphate.
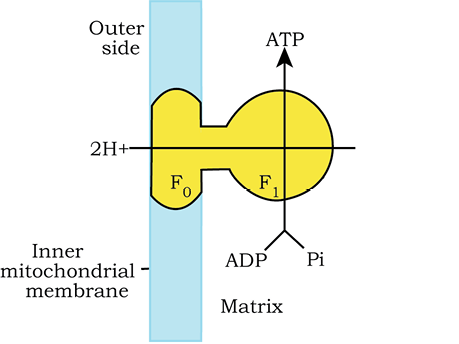
Figure 14.5 Diagramatic presentation of ATP synthesis in mitochondria
F0 is an integral membrane protein complex that forms the channel through which protons cross the inner membrane. The passage of protons through the channel is coupled to the catalytic site of the F1 component for the production of ATP. For each ATP produced, 2H+passes through F0 from the intermembrane space to the matrix down the electrochemical proton gradient.
© 2026 GoodEd Technologies Pvt. Ltd.

