Much before the actual flower is seen on a plant, the decision that the plant is going to flower has taken place. Several hormonal and structural changes are initiated which lead to the differentiation and further development of the floral primordium. Inflorescences are formed which bear the floral buds and then the flowers. In the flower the male and female reproductive structures, the androecium and the gynoecium differentiate and develop. You would recollect that the androecium consists of a whorl of stamens representing the male reproductive organ and the gynoecium represents the female reproductive organ.
Figure 2.2a shows the two parts of a typical stamen – the long and slender stalk called the filament, and the terminal generally bilobed structure called the anther. The proximal end of the filament is attached to the thalamus or the petal of the flower. The number and length of stamens are variable in flowers of different species. If you were to collect a stamen each from ten flowers (each from different species) and arrange them on a slide, you would be able to appreciate the large variation in size seen in nature. Careful observation of each stamen under a dissecting microscope and making neat diagrams would elucidate the range in shape and attachment of anthers in different flowers.
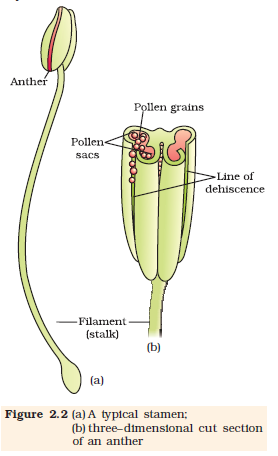
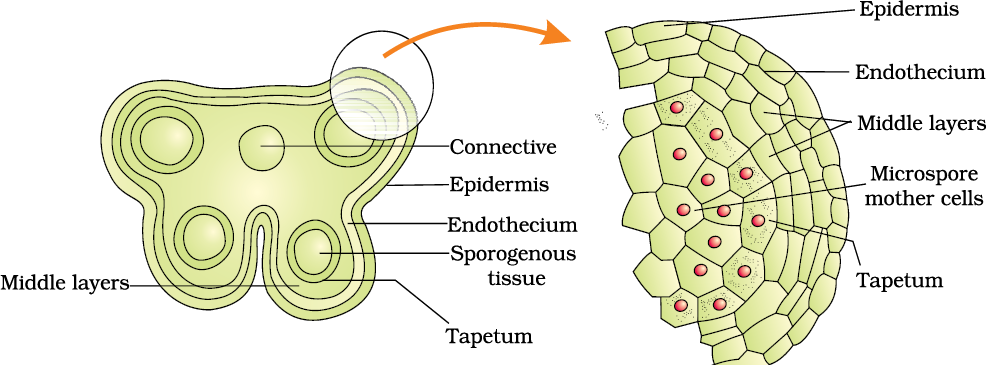
A typical angiosperm anther is bilobed with each lobe having two theca, i.e., they are dithecous (Figure 2.2 b). Often a longitudinal groove runs lengthwise separating the theca. Let us understand the various types of tissues and their organisation in the transverse section of an anther (Figure 2.3 a). The bilobed nature of an anther is very distinct in the transverse section of the anther. The anther is a four-sided (tetragonal) structure consisting of four microsporangia located at the corners, two in each lobe.
The microsporangia develop further and become pollen sacs. They extend longitudinally all through the length of an anther and are packed with pollen grains.
Structure of microsporangium: In a transverse section, a typical microsporangium appears near circular in outline. It is generally surrounded by four wall layers (Figure 2.3 b)– the epidermis, endothecium, middle layers and the tapetum. The outer three wall layers perform the function of protection and help in dehiscence of anther to release the pollen. The innermost wall layer is the tapetum. It nourishes the developing pollen grains. Cells of the tapetum possess dense cytoplasm and generally have more than one nucleus. Can you think of how tapetal cells could become bi-nucleate?
When the anther is young, a group of compactly arranged homogenous cells called the sporogenous tissue occupies the centre of each microsporangium.
Microsporogenesis : As the anther develops, the cells of the sporogenous tissue undergo meiotic divisions to form microspore tetrads. What would be the ploidy of the cells of the tetrad?
(c)
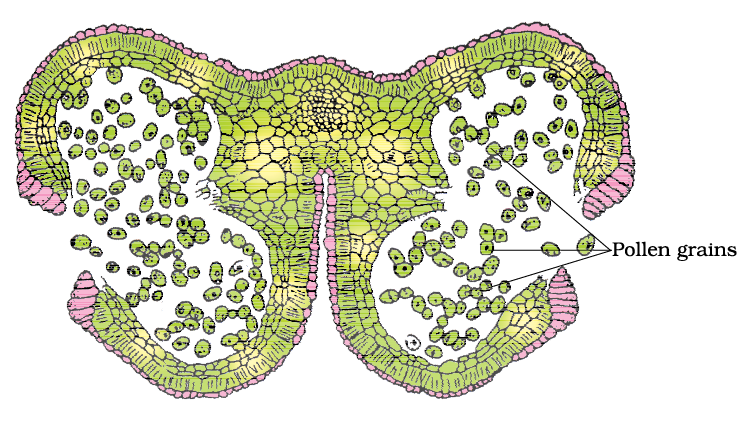
Figure 2.3 (a) Transverse section of a young anther; (b) Enlarged view of one microsporangium showing wall layers; (c) A mature dehisced anther
As each cell of the sporogenous tissue is capable of giving rise to a microspore tetrad. Each one is a potential pollen or microspore mother cell. The process of formation of microspores from a pollen mother cell (PMC) through meiosis is called microsporogenesis. The microspores, as they are formed, are arranged in a cluster of four cells–the microspore tetrad (Figure 2.3 a). As the anthers mature and dehydrate, the microspores dissociate from each other and develop into pollen grains (Figure 2.3 b). Inside each microsporangium several thousands of microspores or pollen grains are formed that are released with the dehiscence of anther
(Figure 2.3 c).
Pollen grain: The pollen grains represent the male gametophytes. If you touch the opened anthers of Hibiscus or any other flower you would find deposition of yellowish powdery pollen grains on your fingers. Sprinkle these grains on a drop of water taken on a glass slide and observe under a microscope. You will really be amazed at the variety of architecture – sizes, shapes, colours, designs – seen on the pollen grains from different species (Figure 2.4).
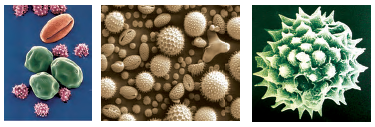
Figure 2.4 Scanning electron micrographs of a few pollen grains
Pollen grains are generally spherical measuring about 25-50 micrometers in diameter. It has a prominent two-layered wall. The hard outer layer called the exine is made up of sporopollenin which is one of the most resistant organic material known. It can withstand high temperatures and strong acids and alkali. No enzyme that degrades sporopollenin is so far known. Pollen grain exine has prominent apertures called germ pores where sporopollenin is absent. Pollen grains are well-preserved as fossils because of the presence of sporopollenin. The exine exhibits a fascinating array of patterns and designs. Why do you think the exine should be hard? What is the function of germ pore? The inner wall of the pollen grain is called the intine. It is a thin and continuous layer made up of cellulose and pectin. The cytoplasm of pollen grain is surrounded by a plasma membrane. When the pollen grain is mature it contains two cells, the vegetative cell and generative cell (Figure 2.5b). The vegetative cell is bigger, has abundant food reserve and a large irregularly shaped nucleus. The generative cell is small and floats in the cytoplasm of the vegetative cell. It is spindle shaped with dense cytoplasm and a nucleus. In over 60 per cent of angiosperms, pollen grains are shed at this 2-celled stage. In the remaining species, the generative cell divides mitotically to give rise to the two male gametes before pollen grains are shed (3-celled stage).
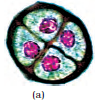
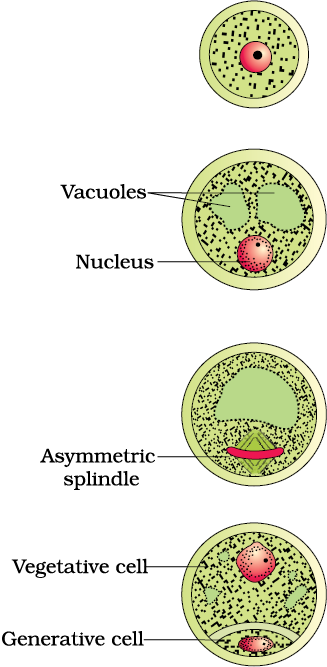
Figure 2.5 (a) Enlarged view of a pollen grain tetrad; (b) stages of a microspore maturing into a pollen grain
Pollen grains of many species cause severe allergies and bronchial afflictions in some people often leading to chronic respiratory
disorders– asthma, bronchitis, etc. It may be mentioned that Parthenium or carrot grass that came into India as a contaminant with imported wheat, has become ubiquitous in occurrence and causes pollen allergy.
Pollen grains are rich in nutrients. It has become a fashion in recent years to use pollen tablets as food supplements. In western countries, a large number of pollen products in the form of tablets and syrups are available in the market. Pollen consumption has been claimed to increase the performance of athletes and race horses (Figure 2.6).

When once they are shed, pollen grains have to land on the stigma before they lose viability if they have to bring about fertilisation. How long do you think the pollen grains retain viability? The period for which pollen grains remain viable is highly variable and to some extent depends on the prevailing temperature and humidity. In some cereals such as rice and wheat, pollen grains lose viability within 30 minutes of their release, and in some members of Rosaceae, Leguminoseae and Solanaceae, they maintain viability for months. You may have heard of storing semen/sperms of many animals including humans for artificial insemination. It is possible to store pollen grains of a large number of species for years in liquid nitrogen (-1960C). Such stored pollen can be used as pollen banks, similar to seed banks, in crop breeding programmes.
The gynoecium represents the female reproductive part of the flower. The gynoecium may consist of a single pistil (monocarpellary) or may have more than one pistil (multicarpellary). When there are more than one, the pistils may be fused together (syncarpous) (Figure 2.7b) or may be free (apocarpous) (Figure 2.7c). Each pistil has three parts (Figure 2.7a), the stigma, style and ovary. The stigma serves as a landing platform for pollen grains.
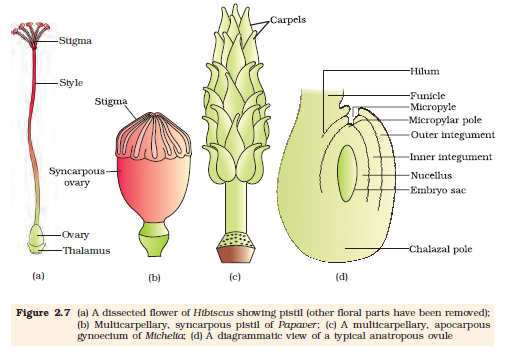
The style is the elongated slender part beneath the stigma. The basal bulged part of the pistil is the ovary. Inside the ovary is the ovarian cavity (locule). The placenta is located inside the ovarian cavity. Recall the definition and types of placentation that you studied in Class XI. Arising from the placenta are the megasporangia, commonly called ovules. The number of ovules in an ovary may be one (wheat, paddy, mango) to many (papaya, water melon, orchids).
The Megasporangium (Ovule) : Let us familiarise ourselves with the structure of a typical angiosperm ovule (Figure 2.7d). The ovule is a small structure attached to the placenta by means of a stalk called funicle. The body of the ovule fuses with funicle in the region called hilum. Thus, hilum represents the junction between ovule and funicle. Each ovule has one or two protective envelopes called integuments. Integuments encircle the nucellus except at the tip where a small opening called the micropyle is organised. Opposite the micropylar end, is the chalaza, representing the basal part of the ovule.
Enclosed within the integuments is a mass of cells called the nucellus. Cells of the nucellus have abundant reserve food materials. Located in the nucellus is the embryo sac or female gametophyte. An ovule generally has a single embryo sac formed from a megaspore.
Megasporogenesis : The process of formation of megaspores from the megaspore mother cell is called megasporogenesis. Ovules generally differentiate a single megaspore mother cell (MMC) in the micropylar region of the nucellus. It is a large cell containing dense cytoplasm and a prominent nucleus. The MMC undergoes meiotic division. What is the importance of the MMC undergoing meiosis? Meiosis results in the production of four megaspores (Figure 2.8a).
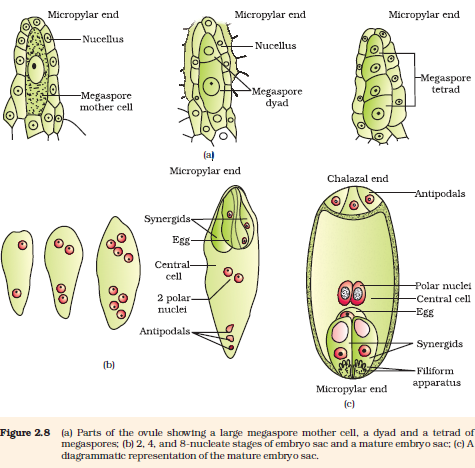
Figure 2.8 (a) Parts of the ovule showing a large megaspore mother cell, a dyad and a tetrad of megaspores; (b) 2, 4, and 8-nucleate stages of embryo sac and a mature embryo sac; (c) A diagrammatic representation of the mature embryo sac.
Female gametophyte : In a majority of flowering plants, one of the megaspores is functional while the other three degenerate. Only the functional megaspore develops into the female gametophyte (embryo sac). This method of embryo sac formation from a single megaspore is termed monosporic development. What will be the ploidy of the cells of the nucellus, MMC, the functional megaspore and female gametophyte?
Let us study formation of the embryo sac in a little more detail. (Figure 2.8b). The nucleus of the functional megaspore divides mitotically to form two nuclei which move to the opposite poles, forming the 2-nucleate embryo sac. Two more sequential mitotic nuclear divisions result in the formation of the 4-nucleate and later the 8-nucleate stages of the embryo sac. It is of interest to note that these mitotic divisions are strictly free nuclear, that is, nuclear divisions are not followed immediately by cell wall formation. After the 8-nucleate stage, cell walls are laid down leading to the organisation of the typical female gametophyte or embryo sac. Observe the distribution of cells inside the embryo sac (Figure 2.8b, c). Six of the eight nuclei are surrounded by cell walls and organised into cells; the remaining two nuclei, called polar nuclei are situated below the egg apparatus in the large central cell.
There is a characteristic distribution of the cells within the embryo sac. Three cells are grouped together at the micropylar end and constitute the egg apparatus. The egg apparatus, in turn, consists of two synergids and one egg cell. The synergids have special cellular thickenings at the micropylar tip called filiform apparatus, which play an important role in guiding the pollen tubes into the synergid. Three cells are at the chalazal end and are called the antipodals. The large central cell, as mentioned earlier, has two polar nuclei. Thus, a typical angiosperm embryo sac, at maturity, though 8-nucleate is 7-celled.
© 2026 GoodEd Technologies Pvt. Ltd.

